
We endorsed Thinking About Evolution as an entry point for dialogue between secular scientists and progressive creationists at Reasons to Believe. These creationists believe the earth is old, but are skeptical about evolution. John Harshman, a secular biologist, took up our invitation to dialogue with them, and writes this response to their book.
The title of the book is Thinking about Evolution, and it is co-written by four progressive creationists at Reasons to Believe. These creationists believe that the earth is ancient, aligned with the mainstream consensus, but they dispute biological evolution. The title is apt, since most of the book focuses on explaining their objections to evolutionary theory.
As a phylogeneticist, I was most interested in the contest between two models, progressive creation and common descent. To my disappointment, there’s precious little about the progressive creation theory. In fact, I’ve been unable to find the progressive creation theory stated in the book. It’s clear that it involves the fiat creation, at various points in earth history, of new “kinds”. But what are those points? What are those kinds? Nowhere is any of that specified. We are left with an undescribed hypothesis whose predictions are unclear and which thus can’t be compared to observations.
One would of course expect any examination of those hypotheses to focus on the main evidence: the nested hierarchy of character data, especially genomic data. But there is none of that. Nested hierarchy is mentioned, once unless I have missed something, very briefly and without any real discussion.
I was forced to fall back on a single chapter, one out of twenty-five, that contains anything related to common descent. Its title is Does Evolution Explain the Fossil Record? The chapter title is unfortunately ambiguous. In evolutionary theory, the term “evolution” refers to descent with modification, including both descent from common ancestors and modifications to lineages during that descent. Descent and the causes of modification are separate questions.
Guided evolution, in which God is the source of some modifications, is an attempted answer to the causes of modification and is compatible with common descent. However, the authors of this book reject guided evolution, for theological rather than scientific reasons, as it turns out. Much of the book is spent attempting to show that natural processes couldn’t produce what we see. But this is quite different from the question of common descent vs. separate creation; even if some features of organisms could not have come about through natural processes, that doesn’t constitute a valid objection to common descent or evidence favoring separate creation. My interest is in comparing those two hypotheses, common descent and separate creation, so we are back to that single chapter, or those portions of it that directly address the question.
So what’s the argument in that chapter against common descent and for progressive creation? While it’s true that the expectation of unguided evolution is that change should be gradual, the expectation of taphonomy and stratigraphy is that we won’t be able to find most of that in the fossil record, and this confusion of the complete history with the preserved fraction of history pervades the chapter. Further, change is “gradual” over human-comprehensible expanses of time (such as decades or even millennia) but such periods are geological instants. Eldredge and Gould, in their idea of punctuated equilibria, envisioned change proceeding by ordinary processes during geologically brief periods of thousands of years. Moreover, the rate of phenotypic change would be episodic, not constant. (Recall also that arguments against unguided evolution are not arguments against common descent; again, those are separate questions.)
Does the fossil record show changes that are too fast for known processes in evolution? In fact, the rates of change that can be estimated from the fossil record are far slower than those observed within modern populations, and this can be accounted for by the episodic nature of evolutionary change.
The author misunderstands punctuated equilibria, tying speciation to geological cataclysms and saltations, which Eldredge and Gould did not.
The author argues that rapid diversification events are contrary to evolutionary expectations. The poster child here is the Cambrian explosion, but unfortunately we are given a straw-man description similar to that in Steven Meyer’s Darwin’s Doubt. All the modern phyla (kinds?) are said to appear in almost the same instant of geological time, without precursors. As the book puts it, “For the first four billion years of Earth’s history (the Precambrian), there was no sign of the animal life we know today. Then, within a short period of geological history, new marine animals representing almost all the body plans existing today appeared. They were fully formed and fully complex.” Almost everything in those three sentences is misleading. (A better account than Meyer’s can be found in the book by Erwin and Valentine, cited below.)
There are Precambrian animal fossils, though of course they aren’t what “we know today”. The period over which extant body plans appeared is unclear for several reasons: “body plan” is notoriously ambiguous; “appeared” conflates actual first existence with appearance in the fossil record. Finally, the Cambrian taxa were not “fully formed”, if by that is meant that they display all the characters shown by modern species (i.e., crown groups). For example, Cambrian vertebrates have no internal bones and are very unlike modern vertebrates.
And what does this mean for progressive creation? Are we to take these “body plans”, possibly intended to be equivalent to phyla, to be kinds, with modern species descended from Cambrian ones? Or are all the species separately created variations on a theme, with God apparently unable to come up with any new themes after the Cambrian? The progressive creation theory remains too undefined to assess scientifically.
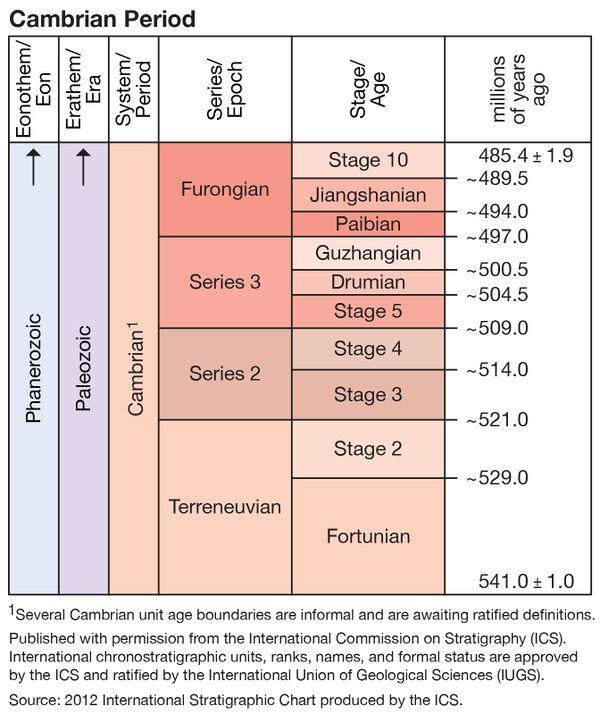
Now in fact there are transitional fossils, some of them Precambrian, some of them Cambrian. To list a couple, the Precambrian Kimberella is clearly some kind of primitive lophotrochozoan, conceivably even a mollusk, though outside any living mollusk group; the Cambrian Anomalocaris is a primitive arthropod, with jointed limbs, though only two and only on its head. It’s unclear just what the author encompasses within the Cambrian explosion. As with Meyer, the Cambrian explosion expands and contracts as convenient to the moment. At one point it’s said that “from the start of the Cambrian, new body plans were so disparate from each other that they were already separate at the phylum level”; and yet most of those body plans are unknown until 20 million years into the Cambrian, at the start of Cambrian Stage 3, in the Chengjiang fauna. Previous to that, most evidence consists of two sorts: trace fossils (tracks and burrows) and the “small, shelly fauna”, both of which begin in the latest Precambrian and gradually increase in diversity and complexity through Cambrian Stages 1 and 2. These, plus a few Precambrian body fossils such as Kimberella, are the precursors the author claims do not exist. The author’s statement neglects around 30 million years of fossil history.

The author displays a serious misunderstanding of taphonomy — the science of fossil preservation — when she claims that if some soft-bodied forms are preserved that shows that no other soft-bodied forms were in existence but unpreserved. In reality there are many sorts of preservation, all differing in their biases toward preserving particular sorts of fossils. The presence of millions of years of trace fossils with no preservation of the actual organisms that made those traces should be instructive.
The author cites the major work on the explosion, Erwin and Valentine 2013, but appears to have read only a single sentence from the introduction. The work is cited in support of the claim that “research seems to indicate that the majority of animal phyla appeared during a period of about 10 million years in the middle of the Cambrian”. (Note that the explosion has here moved from the beginning of the Cambrian, where it was in the previous passage, to its middle, though the author makes no mention of the inconsistency.) The actual sentence says “a great variety and abundance of animal fossils appear in deposits dating from a geologically brief interval between about 530 to 520 Ma, early in the Cambrian period.” That may seem to say the same thing, but the period referred to here is roughly Cambrian Stage 2, a time in which trace and small, shelly fossils gradually increase, though it may extend enough into Stage 3 to encompass the Chengjiang fauna. In context, this is explicitly referring to preservation, not actual origin. And if one reads further into Erwin & Valentine, the Cambrian explosion as described by the authors extends far longer than 10 million years; picking for example Fig. 6.6, the explosion begins (gradually) in Cambrian Stage 1, around 535 Ma, and tapers off (again gradually) after Stage 5 in the Drumian, perhaps 505 Ma. Thus the explosion in that figure encompasses 30 million years and is preceded by a further 20 million years of gradual increase.
A second reference, Lindemann et al. 2019, used to put the start of the explosion at yet another date, 538.5 Ma, also shows gradual accumulation and increase of both trace and small, shelly fossils both before and after that date. See, for example, Fig. 6, and note that the record here shows none of the “body plans” mentioned in the book, only trace fossils, which the book otherwise ignores.
But do those body fossils, when they do arrive in Cambrian Stage 3, appear “fully formed”? They do not. Budd and Jensen (2000) show that almost all Cambrian forms are stem members of their phyla and classes, lacking some of the diagnostic characters of the modern taxa they represent. The various Chengjiang “lobopods” are a case in point, as they contain fossils most closely related to at least three extant phyla — Arthropoda, Onychophora, and Tardigrada — yet without a complete set of diagnostic characters of any of them.
These and other transitional fossils are discounted because they appear for the first time in the Chengjiang fauna at the same time as, for example, crown arthropods, and “Presumed ancestors and descendants should not appear simultaneously with each other”. This conflates both meanings of “appear”, biological and fossil, as well as ancestors and transitional fossils. The first fossil appearances of these taxa are taphonomic artifacts (and in fact some of them are seen earlier, as fragments, in the small, shelly fauna). And the comment about ancestors is a version of “if we came from monkeys, why are there still monkeys?”
The Cambrian explosion is not the only rapid radiation used to cast doubt on common descent. In addition the chapter mentions vascular plants, flying insects, the Triassic recovery from the end-Permian extinction, flowering plants, flowering plants (again), orders of placental mammals, and orders of birds. Only three of these are supported by references, all of them problematic. Most importantly, the use being made of the reference in each case requires acceptance of the phylogeny on which each is based, but paradoxically, each reference is being used to deny that phylogeny’s validity.
First, Soltis et al. 2008 is used to support the statement that “Approximately 97% of all angiosperms make their first appearance within a period of about 5 million years”. This is a serious distortion of the source, which actually says, “After initial, early ’experiments’, angiosperms radiated rapidly (<or=5 million years [Myr]), yielding the five lineages of Mesangiospermae (magnoliids and Chloranthaceae as sisters to a clade of monocots and eudicots + Ceratophyllaceae). This radiation ultimately produced approximately 97% of all angiosperm species.” So in fact it isn’t 97% of angiosperms that appear during this 5 million years, only 5 lineages as broad as monocots and dicots. That 97% comes much, much later. Note also that Soltis et al. base the time span of the 5-million-year radiation on the branch lengths of a molecular phylogeny, so accepting the conclusion also requires accepting the phylogeny and concluding therefore that angiosperms are all related by common descent. The author is thus caught in a paradox, accepting a phylogeny as the foundation of its refutation.
Second, Archibald 2003 (for mammals) and Prum et al. 2015 (for birds) are used to support the claim that “Whole orders of mammals and most orders of modern birds appeared within 15 million years of the last dinosaurs being wiped out”. But Archibald makes no such claim, and in fact finds the data on ordinal appearances ambiguous, concluding “Fossils place the origins of orders around 65mya as do some molecular studies, but others suggest ordinal diversification as old as 100mya.” Further, these orders have precursors in the fossil record.
Prum et al. likewise make no such claim for birds, but an explanation of that requires further digression. If you aren’t interested in a lengthy explanation, here’s the short version: This claim, like that for angiosperms, depends both on a misinterpretation of the source and on the truth of a molecular phylogeny, which truth is then used in another paradoxical attempt to deny its truth. If that’s enough for you, skip the next three paragraphs. But if you’re willing to follow that lengthy explanation, keep reading.
First, note that drawing any conclusions from Prum et al. requires interpreting a time-calibrated molecular tree; more importantly, it requires acceptance of that tree as a valid indicator of phylogeny, as is the case with Soltis et al. Thus birds are one kind and any rapid radiation within them must happen through descent, not creation. Further, the tree doesn’t unambiguously support a claim that most avian orders originated within 15 million years of the K/T boundary.
But understanding that requires even further digression. There are three possible definitions for a taxon: it may begin at the split of a common ancestor between a taxon and its sister group (called a branch-based taxon); it may begin when the common ancestor of all species within the taxon has its first split (called a node-based taxon); or it may begin with the evolution of some diagnostic character (a character-based taxon). The first two may be determined using a phylogenetic tree, but all that can be said of the third is that it must happen some time between the other two. The number of avian orders also varies depending on which classification one goes by. So which definition of “order” does this chapter employ? We don’t know, and how many orders originate in the first 15 million years of the Cenozoic depends on that choice. Under the branch-based definition, of 36 orders in a recent reference (Winkler, Billerman, and Lovette 2015), all but four fall within the 15-million-year window: Struthioniformes (precedes), Caprimulgiformes (precedes), Phoenicopteriformes, Podicipediformes. Under the node-based definition, only seven fall within the window: Struthioniformes, Caprimulgiformes, Suliformes, Pelecaniformes, Coraciiformes, Piciformes, Passeriformes. Seven more can’t be determined, since only one species was sampled (it takes two to make a node) and the corresponding branch-based group is old enough for there to be ambiguity: Otidiformes, Eurypygiformes, Phaethontiformes, Gaviiformes, Sphenisciformes, Opisthocomiformes, Leptosomiformes. Thus at least twenty-two orders do not fall within the window. I should also add that the time calibration used in that tree is controversial, some fossils seem to contradict it, and several relationships hypothesized in that tree are also controversial. The book, of course, must simultaneously claim those relationships both do and do not exist.
Further, what is supposed to have been created in these two radiations? Are kinds to be considered orders? The definition of “kind”, as in the Cambrian explosion, seems flexible enough to encompass every momentary need.
And how are rapid radiations supposed to be signals of progressive creation? Worse, since many of them directly follow from mass extinctions, doesn’t that make God an incompetent who frequently finds himself at an impasse and is forced to wipe the slate clean and start over? How are mass extinctions, or any extinctions for that matter, supposed to fit into a planned, progressive creation? Even discounting mass extinctions, progressive creation implies a God who’s a tinkerer, never able to create more than variations, most of them slight, on a prior theme. Even after mass extinctions, the new creation seems assembled from shards of the previous one.
Why, in fact, is there a limited set of body plans? If phyla are kinds, why is there nested hierarchy among kinds: Lophotrochozoa, Ecdysozoa, Deuterostomia, etc.? If orders are kinds, the same question arises. And if species are kinds, why is there any nested hierarchy at all?
Transitional forms are disposed of by narrowing their definition. In order to count, they must be part of a very gradual and lengthy ancestor-descendant sequence. Whales are used as a case in point. Now of course these are not ancestor-descendant sequences, or at least if they were we would be unable to tell; that’s a limitation of fossil data. But the author additionally tries to claim that they aren’t even intermediates. This is done by ignoring most of the fossils and concentrating only on two, Basilosaurus, clearly a fully aquatic animal, and Maiacetus, which was semi-aquatic, “no more aquatic perhaps than a giant otter”. But what of other fossils mentioned in the text but otherwise ignored, such as Rodhocetus, more reminiscent of a seal than an otter?
Still, the greatest problem with this chapter is that it ignores the bulk of the evidence for common descent, i.e. the nested hierarchy of extant organisms. Perhaps that’s off-topic for a chapter on the fossil record, but this is not addressed elsewhere in the book either. No place else is phylogeny, or anything related to it, even discussed. At best, the chapter is evidence against unguided evolution — though even that is dubious. So why present it as evidence against common descent? Why neglect the evidence favoring common descent?
And just what is the progressive creation model and what are its predictions? As mentioned earlier, the book never actually says. At the conclusion of this chapter, we get “The sudden proliferations of new life forms seem to be more in keeping with the events described in progressive creation models, both in the details provided and in the order in which they are presented.” This is supported by a reference to a web article, Biology’s Big Bangs by Fazala Rana, which merely repeats the same claim at slightly greater length. It contains a link that promises details on the creation model, but that link leads only to “Error 404”. So we are left with a nonexistent model that we are assured is supported by fossil evidence, with no mention that other evidence must also be confronted.
Further, even the chapter on the fossil record implies at various points that kinds are generally phyla (based on the discussion of the Cambrian explosion), orders or perhaps families (based on the discussion of other radiations), or even species (based on the appeal to punctuated equilibria and to gaps in the record).
This book has some value because it shows current thinking among progressive creationists about evolution. Unfortunately, it doesn’t engage our best understanding of evolution, and it doesn’t actually present an alternative hypothesis against which to compare common descent either.
If progressive creation is to have any hope of being tested against common descent, we need more details about what groups are claimed to be related by common descent, and what groups are not. Progressive creationists also need to disentangle the two questions — separate creation vs. common descent and guided vs. natural evolution — or neither can be reasonably addressed.
References
Archibald J.D. Timing and biogeography of the eutherian radiation: fossils and molecules compared. Mol. Phylogen. Evol.2003; 28: 350-359.
Budd G.E., Jensen S. A critical reappraisal of the fossil record of the bilaterian phyla. Biological Reviews 2000; 75:253-295.
Erwin D., Valentine J. The Cambrian explosion: The construction of animal biodiversity. 2013; Springer Nature,
Linnemann U. et al. New high‐resolution age data from the Ediacaran–Cambrian boundary indicate rapid, ecologically driven onset of the Cambrian explosion. Terra Nova 2019; 31:49-58.
Meyer S.C. Darwin’s doubt: The explosive origin of animal life and the case for intelligent design. 2013; Harper One.
Prum R.O. et al. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 2015; 526: 569–573
Roberts A.M., Rana F., Dykes S., Perez M. Thinking about evolution: 25 questions Christians want answered. 2020; RTB Press.
Soltis D.E. et al. Origin and early evolution of angiosperms. Ann N Y Acad Sci. 2008;1133:3-25
Winkler D.W., Billerman S.M., Lovette I.J. Bird families of the world: An invitation to the spectacular diversity of birds. 2015. Lynx Edicions, Barcelona.
Links
- https://peacefulscience.org/articles/rtb-thinking-about-evolution/
- http://www.bio.sdsu.edu/faculty/archibald/Archibald03MPE28p350.pdf
- https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1469-185X.1999.tb00046.x
- https://peacefulscience.org/books/cambrian-explosion/
- https://www.zora.uzh.ch/id/eprint/158482/1/Linnemann_et_al-2018-Terra_Nova.pdf
- https://peacefulscience.org/books/darwins-doubt/
- https://www.researchgate.net/publication/282651425_A_comprehensive_phylogeny_of_birds_Aves_using_targeted_next-generation_DNA_sequencing
- https://peacefulscience.org/books/thinking-about-evolution/
- http://www.amborella.net/Publications/Papers/Soltis%20et%20al%202008-Annals%20of%20the%20New%20York%20Academy%20of%20Sciences.pdf
- https://peacefulscience.org/books/bird-families-world/
Jul 26, 2021
Nov 22, 2021
Apr 18, 2024


