Likely the new director of the NIH, Jay Bhattacharya preaches on the Bible and COVID in 2022. Science Communication
Articles
Likely the new director of the NIH, Jay Bhattacharya preaches on the Bible and COVID in 2022. Science Communication

Reviewing Haarsma’s book, there are indeed “many ways” to reconcile evolution with the doctrine of sin… Ancestry Evolution

Assessing whether the geology of the Grand Canyon is consistent with rapid formation under a young Earth, global flood model.

Assessing whether the geology of the Grand Canyon is consistent with rapid formation under a young Earth, global flood model.

Assessing whether the geology of the Grand Canyon is consistent with rapid formation under a young Earth, global flood model.
Many of us still feel Michael Hesier’s absence. I certainly do, and will for some time come. Theology

Tom Ingebritsen was a biology professor at Iowa State University for 24 years when he found something that changed everything. Confession Resurrection

Revisiting Asa Gray’s understanding of designed evolution for the 21st century

The psychological mechanisms of conflict and dialogue between religion and science

A discussion of various proposals regarding Adam & Eve and how they align with science and Catholic theology
If Dr. Marcus Ross has any scientific objections to my work, let him to take them up directly with me. Science

I went to Vanderbilt specifically to study the social determinants of health, specifically those related to outcomes of Black birthing folks. Science Race

I was not afforded the same opportunities for research as students who went to other institutions. My knowledge and abilities were questioned because I went to a smaller, less well-known HBCU. Science Race

What might the future of eating look like? Only time will tell, but our current eating habits suggest a shift towards a more plant based diet. Science Society
I ultimately learned that Christians do not have to strawman science, and scientists do not have to strawman religion. Evolution Theology

Race, origins, and abuse. A scientist learns a great deal about courage and peacemaking from a week at a Southern Baptist seminary. Science Race Society

A pentecostal scientist explains his common cause with scientists at The Smithsonian, implementing a peaceful pedagogical approach. Evolution Society
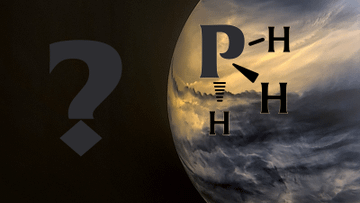
I am one of the scientists studying phosphine in the clouds of Venus. We are still wondering if this is the first signs of life on another planet. Science

John Harshman, a secular biologist, took up our invitation to dialogue with RTB, and he responds to their thoughts on the fossil record. Dialogue Evolution

Drawing on the history of ethics in medicine, perhaps Institutional Review Boards (IRBs) could be how we ensure AI is ethical and good. Artificial Intelligence

Mainstream scientists shouldn’t completely shun organizations friendly to the belief. Reprinted from The Wall Street Journal. Society

The growing conversation around accreditation of creation science, academic freedom, and national norms can be guided by better information. Science Society

It takes real courage for institutions to transparently admit mistakes and to correct them. The ASA offers an olive branch. I accept. Dialogue Society

Let’s play Rob O’Malley’s imagination game. How would you respond? How would you build bridges across these significant divides? Science Society