This excerpt is Chapter 11 of In Quest of the Historical Adam: A Biblical and Scientific Exploration by William Lane Craig, © 2021 by Eerdmans Publishers, Grand Rapids, Michigan. Used by permission. All rights reserved. Figure numbers were altered by removing chapter-prefix, and the title of the chapter was replaced.
Symbolic Behavior
Symbolic thinking is the quintessential human cognitive ability. We earlier appealed to prehistoric art to establish a firm terminus ad quem for the origin of humanity. Now we want to examine various archaeological signatures of such symbolic behavior.
Image and Representation
In recent years, discoveries have pushed the date of prehistoric art into the ever more distant past. On the island of Borneo, Indonesia, a cave painting of a bull has been dated by uranium-thorium testing of calcium carbonate deposits overlying the figure to greater than 40 kya (fig. 1).1

In Sulawesi, Indonesia, the oldest-known instance of figurative art, depicting a scene with a buffalo and pigs, and perhaps human hunters, has been dated by uranium-thorium testing of calcite deposits overlying the figures to a minimum age of 44 kya (fig. 2).2 These caves also contain, as we have seen, hand stencils of comparable age.

Now even older hand stencils have been identified in Maltravieso Cave in Spain, along with other instances of nonfigurative paintings in La Pasiega Cave and Ardales Cave. Uranium-thorium testing of carbonate crusts overlying the stencil dated it to at least 66.7 kya and dated the paintings collectively at a minimum of 64.8 kya, predating the arrival of Homo sapiens in Europe by some twenty thousand years. “The implication is, therefore, that the artists were Neandertals.”3
Reflecting on the significance of this finding, Hoffmann et al. state,
This cave painting activity constitutes a symbolic behavior by definition, and one that is deeply rooted. At Ardales, distinct episodes over a period of more than 25 ka corroborate that we are not dealing with a one-off burst but with a long tradition that may well stretch back to the time of the annular construction found in Bruniquel cave, France, dated to 176.5 ± 2.1 ka ago. Dating results for the excavation site at Cueva de los Aviones, Spain, which place symbolic use of marine shells and mineral pigments by Neandertals at >115 ka ago, further support the antiquity of Neandertal symbolism.4
Given that the use of imagery and representation in art is a signature of modern human behavior among Homo sapiens, it would be prejudicial to deny the humanity of the Neanderthal artists. The contemporaneous presence of similar cave art in both Spain and Indonesia half the world away and the age of ornamental use of shells by Neanderthals imply an origin of symbolic behavior and hence humanity that is vastly older still. Hoffmann et al. conclude, “The corollary of these findings is that the capacity for symbolism must have been inherited from a common ancestor. As a working hypothesis, we suggest that the origins of language and the advanced cognition characteristic of extant humans may precede the period before the divergence of the Neandertal lineage, more than half-a-million years ago.”5
Pigment
Closely related to prehistoric art is the processing and use of pigment. The traces of pigment use in the archaeological record most frequently involve iron oxides, which are found in the form of red hematite or yellow limonite. Whether pigment was used for artistic or decorative purposes or for merely functional purposes such as tanning hides, sunscreen, or medicine can be ambiguous in isolated cases, but as Sally McBrearty and Alison Brooks remind us, “If metallic oxides are recovered in association with undoubted art objects, or at sites whose inhabitants are known to have functioned within a well articulated symbolic system, the materials are usually assumed to have been used as a coloring medium.”6 Such an aesthetic interest is symptomatic of modern humanity.
In the MSA levels of Blombos Cave, South Africa, two ocher pieces bearing similar engraved geometric patterns have been found, dating to 77 kya. Francesco d’Errico comments, “The presence of symbolic engravings on artifactual pigment makes it unlikely that the thousands of pigment fragments found at Middle Stone Age sites were strictly functional and suggests instead that they were used for symbolic purposes.”7 Evidence for “nonfunctional” use of pigment includes the deliberate preference for intense red hues, preference for pigment from faraway sources, deliberate heating to change pigment color, presence of pigment on just one side of an object, coloration of shell beads, and so on.8 Moreover, what we have seen above concerning the antiquity of cave art should settle the question.
Evidence indicates an ancient and widespread use of pigment in Africa during the MSA. Excavations at the site of GnJh-15 in the Kapthurin Formation uncovered more than seventy pieces of red pigment as part of an assemblage of stone artifacts, fragmentary bone, and ostrich eggshell fragments, covered by many meters of volcanic debris dating from 285 kya. Similarly, excavations at Twin Rivers yielded 176 fragments of pigment in layers dated 400–260 kya at the transition from the Acheulean to the MSA.9 “The chronological attribution of the older pigments from Africa (Kapthurin, Twin Rivers) and their association with Lupemban stone tools [dating to around 300 kya] seem to indicate that the use of pigments originated with Homo heidelbergensis or archaic Homo sapiens,” d’Errico explains. “If colorant use is taken as an archeological indication of symbolic behavior, then the origin of these abilities, traditionally attributed to anatomically modern humans, has to be considered more ancient.”10 That conclusion is confirmed by evidence for extremely early use of specularite—a glittery form of hematite useful only for visual display—at Fauresmith, South Africa, from >500 kya.11
Pigment use is not limited to the African MSA. Neanderthals in Europe were using pigment, mostly black but also red, since approximately 300 kya, though systematic use occurs only after 60 kya. Fragments of pigment come from some forty Middle and Upper Palaeolithic sites in Europe, the richest collection being Pech-de-l’Azé I, dated 60–50 kya. Marie Soressi and Francesco d’Errico reject the idea of an exclusively functional use of pigment on ethnographic grounds: “In traditional societies studied by ethnography pigments are always used for symbolic activities. … If the current model is applicable to Neanderthal society, the systematic use of pigments by these societies is a strong argument in favor of their capacity to produce symbolic cultures.”12 Whatever we make of this argument, the cave paintings at Maltravieso, La Pasiega, and Ardales bear witness, as we have seen, to Neanderthal artistic use of pigment >64 kya. Such use supports the multispecies model of human cognitive capacity.
Burials
Burial of the dead is actually better attested in the archaeological record among Neanderthals than among Homo sapiens. There are only three MSA sites where evidence of burial has been found: Nazlet Khater and Taramsa in Egypt, dated respectively to 40 kya and 68 kya, and Border Cave, which appears to be the oldest MSA burial. Sediments overlying the grave are dated to 105 kya. Interestingly, hematite pencils are found throughout the entire MSA sequence, and the human infant (BC 3) burial site is stained through the application of hematite and associated with a perforated Conus shell, which may have been ornamental.
The earliest evidence for burial among Homo sapiens comes not from Africa but from the Levant at the site of Qafzeh. Of the fifteen individuals represented in the cave, at least four appear to have been deliberately buried. Dated to 120–90 kya, these remains are associated with perforated and ocher-stained Glycymeris shells. Deer antlers were buried with the body of one child. Cultural iconic objects seem also associated with the burials at Skhūl, where a man was buried holding the lower jaw of a massive wild boar, and shells similar to those at Qafzeh are found in the Mousterian layers.
Of the fifty-eight known Middle Palaeolithic burial sites in Europe and the Near East, however, thirty-five belong to Neanderthals. D’Errico reports that there is a growing consensus among palaeoanthropologists that Neanderthals buried their dead.13 We have about twenty reasonably complete Neanderthal skeletons out of over five hundred individuals represented. Complete skeletons are so rare that it is likely that these dead were deliberately buried. Apart from intentional burial of these individuals, it is difficult to explain why articulated skeletons are completely lacking in earlier occupation of caves at numerous sites with good preservation of faunal remains in Middle Pleistocene Europe, in the Near East, and in Africa. So there is “general agreement that most if not all relatively complete Neanderthal skeletons were deliberately interred.”14
Neanderthal burial sites in the Levant are at least as old as those of Homo sapiens. The C layer at Tabūn has been dated by thermoluminescence to 160 kya, making the burial of the Neanderthal specimen C1 the oldest in the world. McBrearty and Brooks observe that burial and other special treatments of the dead are a consistent feature of the symbolic life of modern human societies, so the belief that the Neanderthals deliberately buried their dead has been a major factor contributing to an impression of their humanity.15 This impression does not depend, however, on investing the burial procedure with a ritual significance, though it does not exclude such significance. That the corpses of their fellows were treated differently from dead animals suggests that something more was going on than mere housecleaning.16
Language
The nineteenth-century philologist Max Müller declared, “The one great barrier between the brute and man is Language. Man speaks, and no brute has ever uttered a word. Language is the Rubicon, and no brute will dare to cross it.”17 Language may be thought of as a symbolic and freely extensible communication system, and speech as the externalization of language in sound.18 Extensive studies of so-called animal communication systems bear out Müller’s judgment.19 Language involves symbolism, the use of conventional signs to refer to something other than the animals themselves. Animal vocalizations do not exhibit genuine referentiality but at most so-called “functional reference.” Vervet monkey calls, for example, differ depending on whether the perceived predator is a leopard, a snake, or an eagle, but the calls are not words having leopards, snakes, or eagles as referents. Such calls are not conventional but are hardwired by evolutionary conditioning in vervet brains, and the response to the call by other monkeys can be similarly explained as the result of Pavlovian conditioning. Like a fire alarm, the monkey vocalizing the call lacks any intention of referring, for example, to a leopard, and “specific responses to signals can develop with experience based on simple classical conditioning and without drawing on the concept of information, the meaning of calls, or mental representations of a signal’s purported referent in listeners.”20 Since animal signals lack symbolic meaning, questions like “What did that monkey mean by that signal?” or “What was that monkey referring to?” are ill-posed.21 Such calls are at best functionally referential; that is to say, although context-specific calls may function much in the same way as human words like “Fire!” do, that implies nothing about the underlying mental processes involved. Thus, “the vervet leopard alarm does not refer to leopards in the way the English word ‘leopard’ does. Rather, to a listener the call means that a leopard is present in the same way that the leopard’s growl does, or even the way that the sound of dry leaves crunching under a leopard’s foot might.”22 Functional reference is thus wholly different from linguistic reference. Brandon Wheeler and Julia Fischer conclude: “The concept of functional reference, while historically important for the field, has outlived its usefulness and become a red herring in the pursuit of the links between primate communication and human language.”23 Similarly, while chimpanzees in captivity have been trained to punch certain buttons or make a gesture in order to obtain various specific foods, there is no reason to think that when the chimpanzees press a button or select a picture depicting a banana, they are referring to a banana.24 They are conditioned by training to participate in response-reward activity that is at best functionally referential in obtaining the desired benefits. Trained chimpanzees are thus no nearer to genuine language acquisition. As fascinating as the question of how language and its externalization in speech originated may be, it is only indirectly related to the question that interests us—namely, when they originated. Language’s distinctness from animal communication systems at best suggests that language is something that does not originate early in the Homo lineage close to animality. To gain some insight as to the time of its origin, we may look to anatomical, genetic, and archaeological clues.
Anatomical Clues
Anatomically, a large brain size in a hominin is a prerequisite for language capacity, and the presence of a large brain increases the probability of linguistic ability. So Roger Lewin and Robert Foley think that once hominins attained a brain size in excess of one thousand cubic centimeters, there seems to be little doubt that linguistic capabilities existed and that therefore language may have been present at least in Neanderthals.25 “Given their large brains, there is little doubt that Neanderthals are intelligent flexible hominins, even if there may have been some differences between them and modern humans.”26
In addition to brain size, brain organization is vital. Linguistic ability is associated with both Wernicke’s area and Broca’s area, among others, in the brain. Studies of hominin endocasts reveal signs of Broca’s area in Homo rudolfensis and later species but not in Australopithecines.27 Therefore doubt has been cast on linguistic ability among Australopithecines. Unfortunately, as previously mentioned, palaeoneurologists can learn precious little about hominin language ability from fossil endocasts.
Given the paucity of information to be gained from endocasts, investigators have turned to the study of other anatomical features requisite for speech. Hearing will obviously be important for human speech. Bones of the human ear can be compared with those of both living apes and fossil hominins. Although the aural capability of chimpanzees is basically identical to our own, human ears do have a higher sensitivity in the 2–4 kilohertz range, reflecting differences in the ossicles of the middle ear. Key features of certain speech sounds lie within this range.28 Fossil remains from both Neanderthals and Homo heidelbergensis exhibit middle ear ossicles that lie within the range of modern human ears.29
What about the capability of speech? Hauser et al. are skeptical about the presence of language outside Homo sapiens. With respect to the palaeontological evidence, they state, “Recent studies suggest that approximately equal proportions of the horizontal and vertical sectors of the vocal tract are necessary for speech production (Lieberman, 2011). This conformation is present in Homo sapiens alone, as a result of the autapomorphic retraction of its face below the neurocranium. This points to a critical change after divergence from the Neanderthals.”30 With respect to the archaeological evidence, they observe that Neanderthals “failed to leave any unequivocal evidence for the symbolic behavior patterns … that characterize modern, linguistic, human beings.” By way of comparison, “the artifactual record of contemporaneous Middle Stone Age sapiens in Africa after about 100k year ago tells a very different story, a qualitative transformation in behavior that was reflected in the earliest symbolic objects, complex planning, multi-stage technologies, and other anticipations of Cro-Magnon cognitive prowess.” Thus, “archaeological evidence … points to the emergence of a language of thought in early Homo sapiens, replete with symbolic representations that were externalized in iconic form. … Whenever this occurred, present evidence suggests it was after [my emphasis] our divergence with Neanderthals, and thus, a very recent event.”31
Let us look more closely at each of these considerations. Consider first the anatomical features evidenced by palaeontology. Philip Lieberman explains that the position of the larynx is the key to the difference between the supralaryngeal vocal tract (SVT) of all other mammals and the adult modern human SVT.32 The SVT of chimpanzees is significantly different from that of humans, so that although they can hear what we hear, they cannot produce articulate speech.33
In early Homo sapiens there occurred a restructuring of the skull that brought the human face into line with the braincase, thus reducing prognathism. In the process the oral cavity was shortened, forcing the tongue up and back into the throat. The larynx is pushed lower in the throat to a position opposite the fourth, fifth, and sixth cervical vertebrae. The hyoid bone, a U-shaped bone positioned above the larynx and connected to it by ligaments and muscle, also descends with the larynx. The adult human SVT thus features a curved tongue partly occupying the “horizontal” oral cavity and, at a right angle to it, the vertical pharyngeal cavity.
Remarkably, in human ontogeny, from embryogenesis to childhood, this process is recapitulated. The tongue moves back into the pharynx, pushing the larynx down, until the “horizontal” oral cavity and vertical pharyngeal cavity of the SVT have equal 1:1 proportions. In the first two years of life the face retracts and the base of the skull flexes from the relatively flat contour that it had at birth. This cranial base flexion ceases by two to three years of age, but the tongue and larynx continue to descend until ages six to eight, when the proportions of the oral and pharyngeal cavities become equal.
The descent of the larynx in adult humans enlarges the space above the larynx, so that sounds emitted from the larynx can be modified to a greater degree than is possible for any other mammal. Movements of the tongue in the right-angle space defined by the mouth and pharynx are able to produce the changes necessary for utterance of the so-called quantal vowels (phonetically discrete vowels) [i], [u] and [a] (fig. 3).34 By contrast, the tongues of apes, like the tongues of human newborns, are located almost entirely within their mouths, making the production of these vowel sounds impossible.
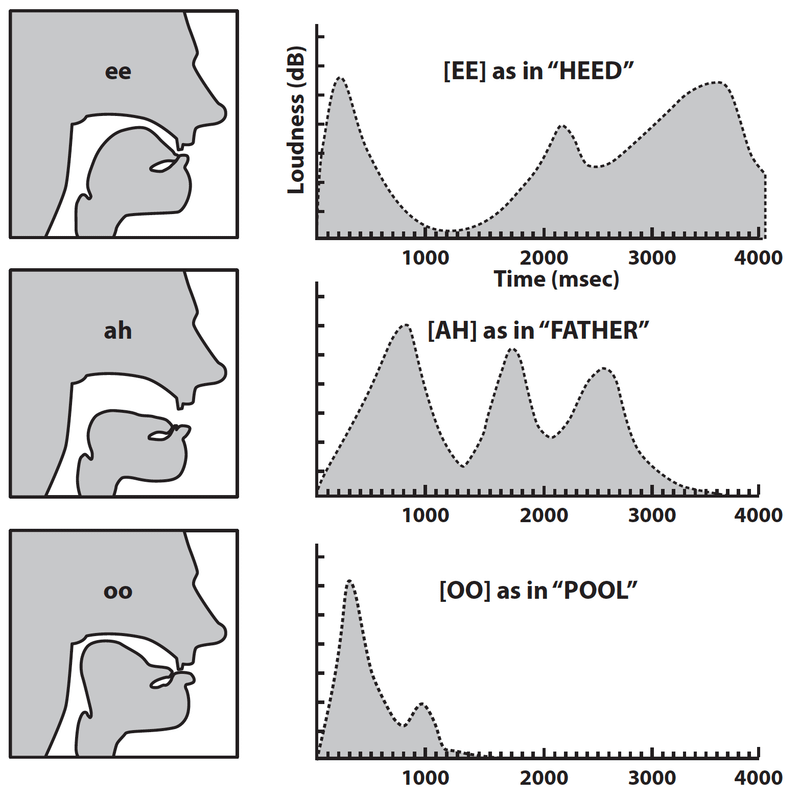
Fossil remains at Skhūl and Qafzeh revealed a fully human SVT in archaic humans 100 kya. Philip Lieberman points out that the biological disadvantages of the human SVT (such as choking on food, impacted molars, reduced chewing efficiency) would reduce fitness unless it was being used to enhance the intelligibility of speech communication.35 The presence of “such an odd, seemingly maladaptive configuration” is thus indicative of articulate speech.36
What about other ancient hominins? The discovery in 1989 at Kebara, Israel, of a Neanderthal hyoid bone virtually identical to the modern bone has been taken by some as evidence for their language ability. Two similar hyoid bones have been found for Homo heidelbergensis. By contrast, a hyoid bone from Australopithecus afarensis is basically apelike. Philip Lieberman protests that it is impossible to determine larynx position and SVT morphology from an isolated hyoid bone.37 Rather, what is critical is the proportion ality of the sections of the SVT. The oral cavity in modern humans measures 57 ± 5.1 millimeters. This is shorter than in Homo heidelbergensis (68.3 ± 5.1 mm) and Homo neanderthalensis (62.3 ± 6.5 mm).38 Moreover, the cranial base of Homo sapiens is approximately 10–15 degrees more flexed, thus retracting the face and shortening the pharyngeal space behind the palette by about one centimeter.39 As a result, the skeletal length of the “horizontal” SVT in ancient Homo sapiens (10.5 cm) is about 10 percent shorter than that of Homo neanderthalensis (11.7 cm) and Homo heidelbergensis (11.8 cm). To have a modern SVT with a ratio of 1:1, they would have needed a vertical SVT two to three centimeters longer than that of the average adult modern human. A modern SVT configuration in an archaic Homo would have positioned the larynx so low that swallowing might become impossible. So Neanderthal vocal tracts could not produce the full range of sounds that characterize human speech, in particular so-called quantal vowels such as [i] in “tea” and [u] in “to.”40 Therefore, the Neanderthal phonetic repertoire was inherently limited.
Lieberman’s claims have been challenged.41 But the more fundamental point is that having an inherently limited phonetic repertoire obviously does not imply a lack of language, for all of us are limited in our phonetic repertoire, even if not inherently so. We may all have the capacity to produce the full range of sounds that characterize human speech in the sense that if we were born and raised in any given culture, we should learn its language without difficulty; but as any American who has struggled to speak a foreign language can testify, there are sounds that native English speakers do not normally use and have difficulty learning, such as the French soeur or the German Mönch. There are living languages that do not, in fact, include any of the quantal vowel sounds.
So Lieberman does not draw the inference that Neanderthals therefore lacked speech. He asks, “Does this mean that Neanderthals lacked speech and language? Probably not. … The archaeological record indicates that they had some form of language and speech.”42 “The general level of Neanderthal culture is such that this limited phonetic ability was probably utilized and that some form of language existed.”43 Indeed, “speech must have been in place in archaic hominids ancestral to humans and Neanderthals. There would have been no selective advantage for retaining mutations that yielded the species-specific human speech producing anatomy at the cost of increased morbidity from choking, unless speech was already present.”44
But he cautions, “Their speech was different, however, being less intelligible than ours.”45 But then we must ask, Less intelligible to whom? Surely Lieberman has chosen the wrong word. Intelligible means “comprehensible.” Are we seriously to think that Neanderthals had difficulty understanding one another? Such a claim not only goes far beyond the evidence but seems quite implausible from what we know of Neanderthal culture. I suspect that Lieberman meant something more like “articulate,” in view of Neanderthals’ limited phonetic range. For he also asserts that “the chimpanzee SVT could produce nasalized, vowel-reduced speech that, though it wouldn’t be as intelligible as normal human speech, would suffice for communication.”46 If such vowel-reduced speech is sufficient for communication, it is necessarily intelligible, even if it is relatively inarticulate. The reason “chimpanzees cannot produce any speech” is that they lack the “specialized brain mechanisms that are necessary to regulate the complex, involuntary, articulatory gestures that underlie speech.”47
Daniel Lieberman explains that what makes the quantal vowels so useful for oral communication is that, being discrete sounds, they are not so easily confused with other vowels. When different speakers utter words in random order, listeners occasionally make identification errors for most vowels—for example, the [ε] in “beg” with the [I] in “big.” But [i] and [u] are not so frequently confused. Since they have almost no overlap among different speakers (see fig. 4), they are especially useful sounds for vocal communication.
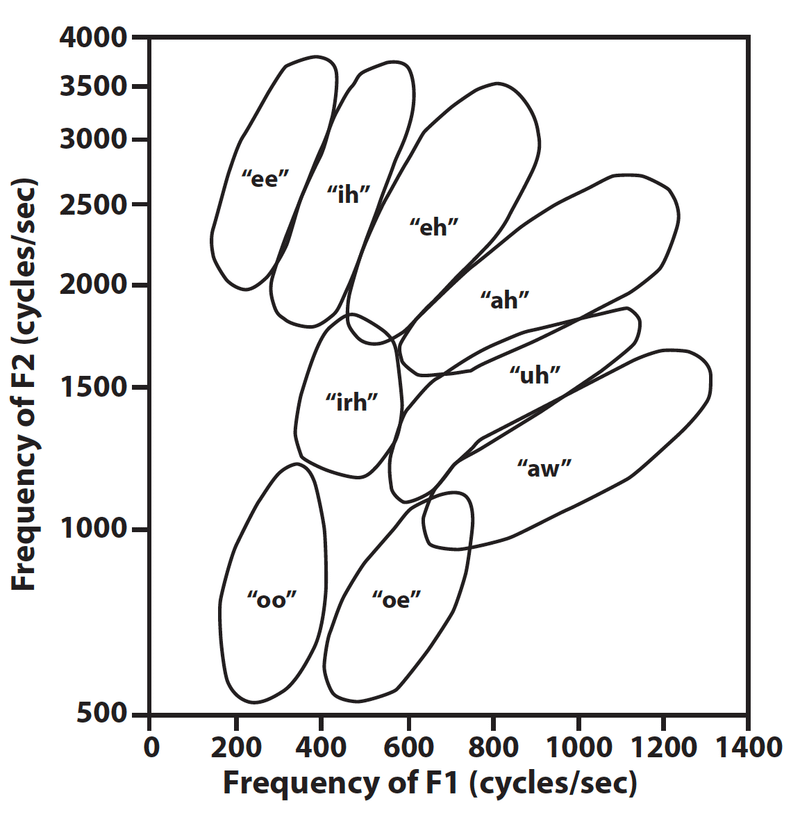
Thus, a speaker can be less precise in articulation and yet still produce sounds such as [i], [a], and [u] with a high degree of perceptibility. “Put differently,” says Daniel Lieberman, “they permit sloppy articulation.”48 Indeed, “one can approximate the vowels with a nonhuman vocal tract of a different configuration, but the formant frequencies are less distinct.”49 Without quantal vowels to set the standard, random vowel sounds would indeed be less intelligible; but then, of course, we are not dealing with random vowel sounds in speech. If we let our imaginations run, we can imagine one Neanderthal saying to another, “That is a beg rhinoceros!” or “I big you to come on the hunt,” and the context of utterance makes the meaning quite plain, despite any problems with articulation. Phonetic differences in articulation need not impinge seriously on the intelligibility of Neanderthal speech to one another.

The speech of little children furnishes a wonderful illustration here. Recall that children do not arrive at an adult configuration of their SVT until ages six to eight, and yet younger children can talk and be understood. Daniel Lieberman observes that children whose SVTs have not yet reached 1:1 proportion “do speak, often well,” but because “their formant frequencies are not as quantal, … perception errors by listeners are higher.”50 But he immediately adds, “Some of these problems, however, may be due to less motor control of the tongue.”51 Of course; and it would be very unfair to expect little children, who are just learning to speak, to exemplify the same degree of articulate speech as an adult Neanderthal, even if the latter also lacked the SVT of a modern adult. Parents of youngsters learning to talk are especially good at interpreting their speech, and we can surmise that Neanderthals would be similarly accustomed to hearing and understanding one another correctly. Noting studies that purport to show that a Neanderthal equipped with a modern SVT would have a larynx impossibly low in the chest, Daniel Lieberman asks, “If true, does this result mean that Neanderthals, other species of archaic Homo, and possibly even some early modern humans couldn’t speak? Of course not. It is hard to imagine that they lacked the capacity for speech, particularly given the large size of their brains. But it may be possible that their articulation was less precise than an adult modern human’s, perhaps more like that of a 4–6 year-old, lacking fully quantal eehs and oohs.”52
As for Hauser et al.’s second point—namely, the lack of archaeological evidence of Neanderthal cognitive capacity—our survey of the archaeological evidence leads to a quite different conclusion, that Neanderthals compared very favorably with MSA Homo sapiens in anticipating Cro-Magnon cognitive ability. I must agree with Dan Dediu and Stephen Levinson that
language affords culture-carrying capacity (e.g. there are no advanced technologies without language), and this linkage allows reasonable inferences from the archeological record. Therefore, we think it is overwhelmingly likely that Neanderthals were as much articulate beings as we ourselves are, that is, with large vocabularies and combinatorial structures that allowed propositional content and illocutionary force to be conveyed. Only such an advanced communication system could have carried the advanced cultural adaptations that Neanderthals exhibited. …
If one considers all of the cultural skills needed to survive in ecologies from the Arctic to game-poor Mediterranean littorals, it is difficult to argue that Neanderthals lacked complex linguistic codes, capable of communicating about spatial locations, hunting and gathering, fauna and flora, social relations, technologies, and so on. This would imply a large lexicon, and propositional encoding. Granting Neanderthals advanced language capacities seems to us inevitable.53
We have seen that the evidence supports a multispecies development of modern cognitive capacity rather than the single species African model espoused by Hauser et al. At the end of the day, Hauser et al. admit, “In terms of the archaeological record, we can certainly imagine the discovery of richer symbolic artifacts … dating before the emergence of Homo sapiens. Such findings would push back the origins of symbolic capacities, and provide greater traction into questions of both origin and subsequent evolution.”54 The artistic representations subsequently discovered at Neanderthal sites in Spain seem to have supplied evidence of just the symbolic artifacts that, if not themselves dating before the emergence of Homo sapiens, disclose a cognitive capacity independent of Homo sapiens, which is therefore just as significant for questions of the origin and subsequent evolution of language.
Are there other anatomical clues to language capacity in Neanderthals? Recall our discussion of the arterial foramina, which are indicative of increased brain metabolism in Neanderthals consistent with the capacity for speech.55 R. F. Kay, M. Cartmill, and M. Balow have also pointed to the hypoglossal canal through which the nerves controlling the tongue pass as indicative of a capacity for speech not only in early Homo sapiens but also in Homo heidelbergensis and Homo neanderthalensis. They found that, by contrast, the hypoglossal canals of Australopithecines, and perhaps also Homo habilis, not only are significantly smaller than those of modern humans but fall within the range of modern chimpanzees, who, it will be recalled, lack the motor control of the tongue and lips requisite for speech. Kay, Cartmill, and Balow conclude, “The vocal abilities of Australopithecus were not advanced significantly over those of chimpanzees whereas those of Homo may have been essentially modern by at least 400,000 years ago.”56
Subsequent studies, however, have challenged their conclusions, finding no correlation between the size of the hypoglossal canal and language ability.57 Monkeys, for example, have no capacity for speech, yet more than half the monkeys measured have hypoglossal canals that are in the modern human size range, both absolutely and relative to mouth size. These results show that a large hypoglossal canal is no guarantee of linguistic ability. But that was surely not Kay, Cartmill, and Balow’s claim. In addition to motor control of the tongue, a large brain is a prerequisite for language. Their claim is better understood to be that a large hominid brain and a large hypoglossal canal are jointly sufficient for, or at least evidence of, language ability.58 To refute this claim one would need to point to language users who have a large brain but a narrow hypoglossal canal, which has not been done. At the least, the large hypoglossal canal in various Homo species is consistent with their being capable of speech.
Additional anatomical evidence comes from the enlarged thoracic vertebral canal in human beings, through which pass the nerves that control the muscles used to regulate breathing. Such fine respiratory control is crucial for human speech. Ann MacLarnon and Gwen Hewitt explain that “full human language requires extended exhalations for vocalizations and increased control of volume, emphasis, and intonation compared with nonhuman primates and therefore presumably compared with early hominids. Such features require fast, intricate, flexible, and integrated neural control of intercostal and abdominal muscles.”59 The muscles enabling such fine respiratory control are all thoracically innervated.
So MacLarnon and Hewitt measured the thoracic vertebral canal for a variety of fossil hominins: Australopithecus afarensis, Australopithecus africanus, Homo ergaster (or early Homo erectus), four Neanderthals, and one early Homo sapiens, in addition to three modern human samples. They found that both the Neanderthals and the early and contemporary modern humans have larger relative thoracic canal cross-sectional areas than any of the non-human primates. “This evidence indicates that thoracic innervation in earlier fossil hominids, australopithecines and Homo ergaster, was similar to that of extant nonhuman primates, but that Neanderthals and early modern humans had expanded thoracic innervation similar to that in extant humans.”60
Examining the possible explanations for this increased thoracic innervation—such as postural control for bipedalism, increased difficulty of parturition, respiration for endurance running, an aquatic phase, choking avoidance, and increased control of breathing for speech—MacLarnon and Hewitt argue that all but the last can be ruled out, either because of their evolutionary timing or because they are insufficiently demanding neurologically. By way of positive evidence for this explanation, they appeal to studies of human speech production, which show that human speech “requires very fast, fine control of subglottal pressure which responds to cognitive factors and is integrated with control of the upper respiratory tract and other body changes,”61 and to comparisons of human speech production and the vocalizations of nonhuman primates, which show that human speech is unique in the respiratory control required for its production. MacLarnon and Hewitt conclude, “Neanderthals and early modern humans had expanded thoracic innervation, like extant humans. It seems most probable that this increased innervation evolved to enable enhanced breath control, and the most likely functional reason for this was the evolution of human speech, i.e., the physical production of language.”62
Even if one does not accept that the explanation for increased thoracic innervation is the need of respiratory control for speech, minimally what MacLarnon and Hewitt’s results imply is that one of the singly necessary and jointly sufficient conditions for human speech is already present, along with other similar conditions, in Neanderthals. Neanderthal speech is thus more probable given the presence of this condition than otherwise, so that it constitutes evidence for Neanderthal speech.
Genetic Clues
The sequencing of a complete Neanderthal genome in 2010 revolutionized palaeoanthropological studies. We now have nearly twenty partial or complete specimens of the Neanderthal genome, which can be compared to the genome of modern humans. As a result, the debate as to whether Neanderthals and Homo sapiens interbred has now been decisively answered in the affirmative on the basis of evidence of mutual introgression of genetic materials (fig. 5). Approximately 2 percent of the DNA of all living nonAfricans is derived from Neanderthals, and Oceanic populations have an additional 2–4 percent of their DNA from Denisovans.
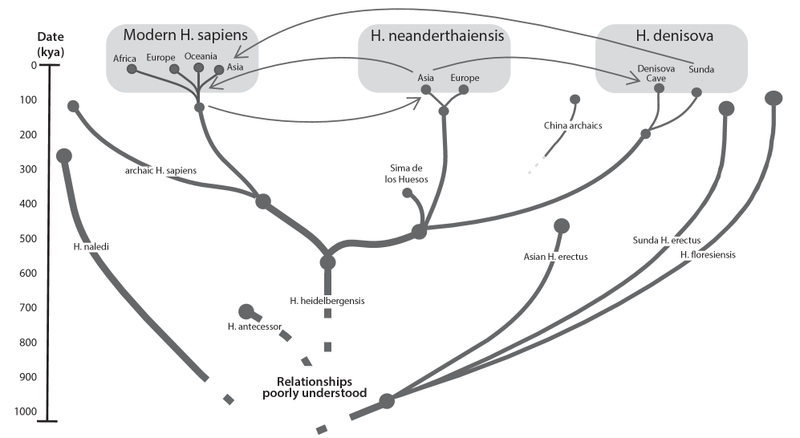
The simple fact alone of interbreeding carries implications for the capacity of Neanderthals and Denisovans for speech. They interacted repeatedly with anatomically modern human beings over tens of thousands of years, in Dediu and Levinson’s memorable words, “exchanging genes, parasites and culture.”63 Such social and sexual intercourse plausibly requires communication and hence language. Kai Whiting et al. comment, “It seems quite unreasonable to assume that all sexual encounters between the different Homo species were of the non-consensual variety. It is much more likely that at least some of the instances of interbreeding between co-existing Homo species, including anatomically modern humans, were the result of communication and a degree of affection or appreciation. Regardless of the exact dynamics of sexual relations, we know for certain that some resulted in offspring that could claim kinship to more than one set of human species.”64 Thus, interbreeding and the production of hybrid offspring who could, like the Homo sapiens parent, speak are themselves suggestive of language ability in the partners.
We have already mentioned crucial mutations in the gene ARHGAP11B and the NOTCH2NL genes in the human lineage prior to the divergence of Homo sapiens, Neanderthals, and Denisovans, and hence shared by all, that contributed significantly to brain growth.65 We do not know if there was any direct effect on language ability as a result.
The significance of the differences between the genome of Homo sapiens and those of Neanderthals and Denisovans remains poorly understood. André Sousa et al. examined so-called human accelerated regions (HARs) of the modern human genome to compare them with genomic material from these ancient hominins.66 HARs are DNA sequences that changed very little throughout mammalian evolution but then experienced a burst of changes in hominins since divergence from chimpanzees. Sousa et al. determined that 8 percent of HAR substitutions are not found in Neanderthals and Denisovans and are thus recent in the sense that the derived allele had not come to fixation in the common ancestor of modern humans and archaic hominins. Some of these substitutions are found in an HAR of AUTS2, a gene associated with several neurological features, in a region also showing strong evidence of a selective sweep that occurred in modern humans after the split with Neanderthals. AUTS2 contains the most significantly accelerated genomic region differentiating humans from Neanderthals, but unfortunately the function and regulation of this gene remain largely unknown.
The celebrated FOXP2 gene is, however, more directly related to a capacity for speech.67 Mutations of this gene have been linked to verbal dyspraxia. So FOXP2 seems to be necessary for human speech. Significantly, human FOXP2, while differing from that of chimpanzees and gorillas in two amino acid coding positions, was found to be identical in Homo sapiens and Neanderthals.68 Neanderthal and Homo sapiens FOXP2 is identical in the very two positions (911 and 977 in exon 7) that differ from ape FOXP2. In this respect, at least, Neanderthals and Homo sapiens would share whatever linguistic benefit these changes confer. Johannes Krause et al. deem the most probable explanation for these shared genetic changes to be that these changes took place in the common ancestor of modern humans and Neanderthals before their divergence.69
Summary
In the last two chapters we have briefly surveyed evidence from palaeoneurology, archaeology, and genetics for the time of human origins.
With respect to the evidence of palaeoneurology, we saw that on the basis of analysis of cranial endocasts there has been in the course of hominin evolution a striking increase in both brain size and encephalization quotient, as well as brain reorganization, that brings hominin brains well into the modern range by 500 kya with Homo heidelbergensis. These features are correlated securely with greater cognitive capacity in humans. Moreover, measurements of the arterial foramina, through which the carotid arteries supplying the brain pass, show that only with later Homo erectus and Homo heidelbergensis do we arrive at the range of modern humans. Larger arterial openings in the skull indicate a higher metabolic rate of the brain, which is suggestive of increased cognitive capacity. Studies of fossil hominin teeth show slow enamel growth in both Neanderthals and archaic Homo sapiens, an index of slow maturation of the brain past birth and well into childhood. The slow trajectory of enamel growth typical of modern humans is not found in Australopithecines or early Homo, which are more apelike in development. Thus, slow brain development is positively correlated with the increased cognitive capacity of modern humans. Genetic comparisons of hominin DNA reveal that a mutation in the gene ARHGAP11B that contributed to the expansion of the human neocortex is shared by both Neanderthals and Denisovans. Since the identical mutation is unlikely to have occurred thrice, it is doubtless a derived feature that was inherited from their last common ancestor, Homo heidelbergensis. Again, the presence of the identical NOTCH2NL genes in Neanderthals, Denisovans, and Homo sapiens points to genomic reorganization events in the brain of their last common ancestor, which resulted in amplification of neuron progenitors and, hence, increased cognitive capacity.
With respect to the evidence of archaeology, the most important evidence for the cognitive capacity of ancient hominins, we saw that generally accepted sufficient conditions of modern humanity may be discerned by the presence of a wide variety of archaeological signatures. These mutually reinforce one another and provide a powerful cumulative case for modern human consciousness that is stronger than its weakest link. We surveyed briefly some of the most important evidence from archaeological signatures in the areas of technology, economy and social organization, and symbolic behavior.
With respect to technology, the production of stone blades, a feature of the Mode 4 toolmaking industry, was practiced by both Neanderthals and Homo sapiens well before 300 kya, marking a technological advance that required significant cognitive capacity to execute. Even more sophisticated was the production of stone points, which both Neanderthals and Homo sapiens crafted at least 186 kya, and which may have been manufactured and employed by Homo heidelbergensis 500 kya. The production of composite tools and hafting required not merely forethought but design and characterized the tool industries of both Neanderthals and Homo sapiens. The extraordinary finds at Schöningen show that 400 kya composite tools were already in use, pointing once more to Homo heidelbergensis. Grindstones are important signatures of cognitive capacity, since their use indicates the processing of plant material and, most significantly, of pigment, one of the signatures of symbolic behavior. They have been found both at MSA sites and at Mousterian sites, indicating use by both Homo sapiens in Africa and Neanderthals in Europe.
With respect to economy and social organization, we saw that big-game hunting involves cooperative behavior indicative of human consciousness and plausibly even language ability. Such behavior evinces a collective intentionality that many psychologists take to be a cognitive skill unique to, even definitive for, human beings.70 Both MSA Homo sapiens and Neanderthals engaged in such hunting activities. Again, the stunning Schöningen spears, whose manufacture alone requires extraordinary cognitive capacity, together with the evidence from Boxgrove and Clacton, show that such behavior goes back to Homo heidelbergensis 500 kya. Equally amazing are the Neanderthal constructions in Bruniquel Cave dating to 176 kya. Nothing like these, involving a chaîne opératoire of astounding complexity and depth, has ever been found before. These constructions exhibit even more clearly than big-game hunting the collective intentionality of their Neanderthal builders. To add to our astonishment, we find the hunters’ huts at Terra Amata, which evince so patently the planning and design of their fabricators. Dating to 350 kya, Homo heidelbergensis was likely responsible for them.
Finally, with respect to the evidence of symbolic behavior, the date of imagistic and representational art among Homo sapiens has now been driven back to >40 kya by the discovery of the Indonesian cave art, and among Neanderthals >66 kya by the Iberian cave art. The contemporaneous presence of similar cave art in Spain and in Indonesia, half the world away, implies an origin of symbolic behavior and hence humanity that is vastly older still. We also saw that the use of pigment, which may be used for art or body decoration, has been attested in Africa to >300 kya and among Neanderthals in Europe to >60 kya. Burials of the dead, whether invested with spiritual significance or not, exhibit a care for the remains of one’s fellows that shows an estimate of their worth. At Qafzeh 120 kya we have the earliest evidence for burial of the dead among Homo sapiens, replete with items interred with the deceased, and at Tabūn 160 kya the earliest evidence for Neanderthal burials. Finally, language use, the paradigmatic symbolic behavior, though difficult to detect, is supported among both Homo sapiens and Neanderthals by anatomical clues like a large and complex brain, aural structures suited to human speech, an SVT suitable to the production of speech despite the dangers posed thereby, enlarged arterial foramina, a large hypoglossal canal, and an enlarged thoracic vertebral canal, as well as genetic clues pointing to interbreeding among Neanderthals, Denisovans, and Homo sapiens and to their sharing the mutated FOXP2 gene crucial to speech that they probably derived from their common ancestor, Homo heidelbergensis.
Accordingly, we have very powerful evidence that human behaviors that exhibit modern cognitive capacity did not originate recently, or even very early, among Homo sapiens alone but were already in place in our last common ancestor with Neanderthals and Denisovans.
References
Craig, William Lane. In Quest of the Historical Adam: A Biblical and Scientific Exploration. (Grand Rapids, MI: Eerdmans Publishers, 2021).
Aubert, M., et al., “Paleolithic Cave Art in Borneo,” Nature 564, no. 7735 (November 7, 2018): 254–57, https://doi.org/10.1038/s41586-018-0679-9.
Callaway, Ewen. “Is This Cave Painting Humanity’s Oldest Story?,” Nature, December 11, 2019, https://doi.org/10.1038/d41586-019-03826-4.
Hoffmann, Dirk. L., et al., “U-Th Dating of Carbonate Crusts Reveals Neandertal Origin of Iberian Cave Art,” Science 359, no. 6378 (February 23, 2018): 912–15, https://doi.org/10.1126/science.aap7778.
Hoffmann, Dirk L., et al., “Symbolic Use of Marine Shells and Mineral Pigments by Iberian Neandertals 115,000 Years Ago,” Science Advances 4, no. 2 (February 2018): eaar5255, https://doi.org/10.1126/sciadv.aar5255.
McBrearty, Sally and Brooks, Alison S. “The Revolution That Wasn’t: A New Interpretation of the Origin of Modern Human Behavior,” Journal of Human Evolution 39, no. 5 (November 2000): 524, https://doi.org/10.1006/jhev.2000.0435.
d’Errico, Francesco. “The Invisible Frontier: A Multiple Species Model for the Origin of Behavioral Modernity,” Evolutionary Anthropology 12, no. 4 (August 5, 2003): 188, https://doi.org/10.1002/evan.10113.
d’Errico, Francesco and Stringer, Chris B. “Evolution, Revolution or Saltation Scenario for the Emergence of Modern Cultures?,” Philosophical Transactions of the Royal Society B 366, no. 1567 (April 12, 2011): 1066, https://doi.org/10.1098/rstb.2010.0340.
Barham, Lawrence S. “Possible Early Pigment Use in South-Central Africa,” Current Anthropology 39, no. 5 (1998): 703–10, https://doi.org/10.1086/204793.
Watts, Ian, Chazan, Michael, and Wilkins, Jayne. “Early Evidence for Brilliant Ritualized Display: Specularite Use in the Northern Cape (South Africa) between ~500 and ~300 Ka,” Current Anthropology 57, no. 3 ( June 2, 2016): 287–301, https://doi.org/10.1086/686484.
Soressi, Marie and d’Errico, Francesco. “Pigments, gravures, parures: Les comportements symboliques controversés des Néandertaliens,” in Les Néandertaliens: Biologie et cultures, ed. Bernard Vandermeersch and Bruno Maureille (Paris: Éditions du CTHS, 2007).
Stringer, Chris and Andrews, Peter. The Complete World of Human Evolution, 2nd ed. (New York: Thames & Hudson, 2012).
Bickerton, Derek. Adam’s Tongue: How Humans Made Language, How Language Made Humans (New York: Hill & Wang, 2009).
Tomasello, Michael. Becoming Human: A Theory of Ontogeny (Cambridge, MA: Belknap Press of Harvard University Press, 2019).
Laland, Kevin N. Darwin’s Unfinished Symphony: How Culture Made the Human Mind (Princeton: Princeton University Press, 2017).
“Helen Keller—Water Scene from ‘The Miracle Worker,’” YouTube video, 6:11, posted by Helen Keller Channel, March 26, 2010, accessed February 2, 2022, https://www.youtube.com/watch?v=lUV65sV8nu0.
Båve, Arvid. “A Deflationary Theory of Reference,” Synthèse 169 (2009): 51–73, https://doi.org/10.1007/s11229-008-9336-4.
Johansson, Sverker. “Language Abilities in Neanderthals,” Annual Review of Linguistics 1 (2015): 313, https://doi.org/10.1146/annurev-linguist-030514-124945.
Hauser, Marc D. The Evolution of Communication (Cambridge, MA: MIT Press, 1996).
Hauser, M. D. et al., “The Mystery of Language Evolution,” Frontiers in Psychology 5 (May 7, 2014), 2-5, https://doi.org/10.3389/fpsyg.2014.00401.
Wheeler, Brandon C. and Fischer, Julia. “Functionally Referential Signals: A Promising Paradigm Whose Time Has Passed,” Evolutionary Anthropology 21, no. 5 (September 2012): 199, https://doi.org/10.1002/evan.21319.
Wheeler, Brandon C. and Fischer, Julia. “The Blurred Boundaries of Functional Reference: A Response to Scarantino & Clay,” Animal Behaviour 100 (2015): e9–e13, https://doi.org/10.1016/j.anbehav.2014.11.007.
Bonnette, Dennis. Origin of the Human Species, VIBS 106 (Amsterdam: Rodopi, 2001).
Lewin, Roger and Foley, Robert A. Principles of Human Evolution, 2nd ed. (Oxford: Blackwell, 2004).
Lieberman, Daniel E. The Evolution of the Human Head (Cambridge, MA: Harvard University Press, 2011).
Tattersall, Ian. The Fossil Trail: How We Know What We Think We Know about Human Evolution, 2nd ed. (Oxford: Oxford University Press, 2009).
Lieberman, Philip. “Current Views on Neanderthal Speech Capabilities: A Reply to Boe et al. (2002),” Journal of Phonetics 35, no. 4 (2007): 552–63, https://doi.org/10.1016/j.wocn.2005.07.002.
Lieberman, Philip. “On Neanderthal Speech and Human Evolution,” Behavioral and Brain Sciences 19, no. 1 (1996): 157, https://doi.org/10.1017/S0140525X00042047.
D’Anastasio, Ruggero, et al., “Micro-Biomechanics of the Kebara 2 Hyoid and Its Implications for Speech in Neanderthals,” PLoS ONE 8, no. 12 (2013), e82261, https://doi.org/10.1371/journal.pone.0082261.
Lieberman, Philip and Crelin, Edmund S. “On the Speech of Neanderthal Man,” Linguistic Inquiry 11, no. 2 (1971): 213, https://www.jstor.org/stable/4177625.
Boë, Louis-Jean et al., “The Potential Neandertal Vowel Space Was as Large as That of Modern Humans,” Journal of Phonetics 30, no. 3 (2002): 481–82, https://doi.org/10.1006/jpho.2002.0170).
Boë, Louis-Jean et al., “Anatomy and Control of the Developing Human Vocal Tract: A Response to Lieberman,” Journal of Phonetics 41, no. 5 (2013): 379–92, https://doi.org/10.1016/j.wocn.2013.04.001.
Lieberman, Philip. “Vocal Tract Anatomy and the Neural Bases of Talking,” Journal of Phonetics 40, no. 4 (July 2012): 613, https://doi.org/10.1016/j.wocn.2012.04.001.
Dediu, Dan and Levinson, Stephen C. “Neanderthal Language Revisited: Not Only Us,” Current Opinion in Behavioral Sciences 21 (2018): 52–53, https://doi.org/10.1016/j.cobeha.2018.01.001.
Kay, R. F., Cartmill, M., and Balow, M. “The Hypoglossal Canal and the Origin of Human Vocal Behavior,” Proceedings of the National Academy of Sciences of the United States of America 95, no. 9 (April 28, 1998): 5417–19, https://doi.org/10.1073/pnas.95.9.5417
DeGusta, David, Gilbert, W. Henry, and Turner, Scott P. “Hypoglossal Canal Size and Hominid Speech,” Proceedings of the National Academy of Sciences of the United States of America 96, no. 4 (February 16, 1999): 1800–804, https://doi.org/10.1073/pnas.96.4.1800.
MacLarnon, A. M. and Hewitt, G. P. “The Evolution of Human Speech: The Role of Enhanced Breathing Control,” American Journal of Physical Anthropology 109, no. 3 (1999): 358. https://doi.org/b2b7q2
Whiting, Kai et al., “Were Neanderthals Rational? A Stoic Approach,” Humanities 7, no. 2 (2018): 39, https://doi.org/10.3390/h7020039.
Sousa, A. M. M. et al., “Evolution of the Human Nervous System Function, Structure, and Development,” Cell 170, no. 2 ( July 13, 2017): 226–40, https://doi.org/10.1016/j.cell.2017.06.036.
Fisher, Simon E. “Evolution of Language: Lessons from the Genome,” Psychonomic Bulletin & Review 24, no. 1 (2017): 34–40, https://doi.org/10.3758/s13423-016-1112-8.
Krause, Johannes et al., “The Derived _FOXP2 Variant of Modern Humans Was Shared with Neandertals,” Current Biology 17, no. 21 (November 6, 2007): 1908–12, https://doi.org/10.1016/j.cub.2007.10.008.
Tomasello, Michael. A Natural History of Human Thinking (Cambridge, MA: Harvard University Press, 2014)
Mar 9, 2022
Sep 28, 2021
Jul 24, 2026